Izadi Jakintza»Izadi jakintza
Mendelen ondorengo genetika klasikoa I
LABURPENA: Beren loci-ak kromosoma berean kokatuak dituzten geneek ez dute betetzen geneen independentziaren Mendelen legea. Aitzitik, gene lotuak dira, berez, belaunaldiz belaunaldi elkarturik transmititu behar direnak. Herentziaren kromosoma teoriaren euskarri nagusietako bat izan zen loturaren fenomeno honen aurkikuntza, lotura multzoen eta kromosomen arteko paralelismoarena. Geroago ikusi zen lotura hori ez zela erabatekoa; izan ere, kromosoma homologoek geneak truka ditzakete eta konbinazio berriak sortu, meiosian kiasmak sortuz, era horretan trukegurutzaketaren edo gene trukearen fenomenoa gertazen dela. Locien artean gertatzen diren gurutzaketen kopurua locien arteko distantzien neurria da, eta, hortaz, neurri hori erabil daiteke kromosomen mapa genetikoak egiteko.
Geneen lotura
Mendel 1900ean berraurkitu ondorentxoan, kasu askotan ikusi ahal izan zen geneen independentziaren legea ez zela betetzen, eta F ren fenotipo proportzio ezaguna 2 (9:3.3:1) beraz ezin zitzaien fenotipo guztiei aplikatu, ezta F bati dagokion 1.1.1.1. atze- 1 rako gurutzaketa ere.
Batesonek eta Punnetek, mende honen hasieran, deskribatu zuten independentziaren legeari jarraitzen ez dioten lehen kasu hauetako bat. Ilar usaintsuaren bi ezaugarriren transmisioa aztertzerakoan, loreen kolorearena eta polenaren formarena alegia, ikusi zuten bi F gurutzatu ondoren atera- 1 tzen zen F a ez zitzaiola egokitzen Mendelen 2 proportzio ezagunari. Aitzitik, P belaunaldian elkarturik zeuden ezaugarrien formek F ra elkarrekin igarotzeko joera zuten.
2 Fenomeno berbera aurkitu zen animalietan.
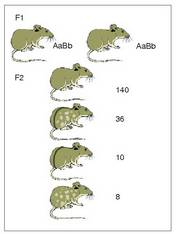
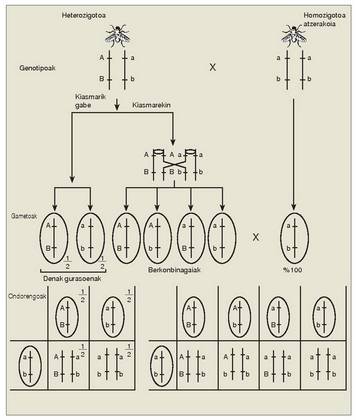
Ikus dezagun adibide bat oso ezaguna, saguena; bi ezaugarriren herentzia aztertu zen: ilearen kolorearen uniformetasuna (kolore uniformea da gainartzailea, eta arrasto zurikoa atzerakoia), eta ilea izan ala ez izan (ile normala gainartzailea, eta soilunekikoa atzerakoia).
Esperimentu batean, F heterozigoto 1 bikoitzen artean hainbat gurutzaketa egin ziren (P gurasoetako bat homozigoto atzerakoia zen denetan, eta homozigoto gainar tzailea bestea); kolorez uniformeak ziren eta ile kopuruz normalak, eta horiek homozigoto atzerakoi arrastodun eta soilunekikoekin gurutzatu ziren. Guztira 194 F lortu ziren, 2 eta hau zen fenotipoen proportzioa:Ikusten denez, forma gehienak gurasoenak
dira, eta gutxienak forma berriak edo berkonbinagaiak
dira. Geneek jokabide hori
dutenean, Mendelen legearen arabera espero
ez zena alegia, lotuak daudela esaten da.
Zein da lotura horren zergatia? Herentziaren
kromosoma teorian dago horren arrazoia.
Saguaren bi geneek kromosoma berean
dute beren locia. Hortaz, meiosian, bikotearen
kromosoma bakoitza osorik joaten da
gametoetako batera, bi alelo dominatzaileak
nahiz bi atzerakoiak berekin daramatzala.
Alabaina, kromosoma bakoitzean bi gene
baino gehiago izaten da. Horren ondorioz,
aurreko azalpena orokorragoa egin daiteke,
eta esan daiteke kromosoma baten loci guztiak
elkartuak daudela, eta beren aleloak
batera igaroko direla belaunaldiz belaunaldi.
Kromosoma berean dauden gene hauei
buruz, esaten da lotuak daudela (elkartuak),
eta lotura talde bat osatzen dutela. Mota
batean, zenbat kromosoma desberdin, hainbat
lotura talde izango dira. Hala, giza
motan esaten da 23 lotura talde direla (gizonezkoan
24 dira).
Horren ondorioz, kromosoma desberdinetan
dauden geneek bakarrik betetzen dute
independentziaren legea. Gainerako egoeretan
lotura dela esan behar da. Nolanahi ere,
ez dirudi kromosoma baten alelo guztiak
ondorengoetara denak batera igarotzen direlako
loturaren hastapena betetzen denik ere.
Nola agertu, bestela, aurreko F an sagu ba-
2
tzuk fenotipo berkonbinagaiekin atera
izana? Lotura bete balitz gurasoen konbinazioak
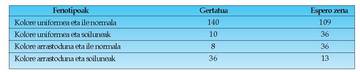
besterik ez ziren aterako, 3:1 konbinazioan. Arazo horren erantzuna hurrengo atalean
dago.Mendelen arabera espero zen emaitza, hau
da, lau fenotipoen ratioa 9.3.3.1 izatea, ez da
betetzen hemen. Horren ordez, F
2
ek 3:1eko
ratioa beharko lukete izan gurasoen fenotipoen
artean, alegia, bi monohibrido gurutzatzean
lortzen den bera. Beste bi fenotipo
posibleak, berkonbinagai deritzaienak, ez
lirateke agertu behar.

Trukegurutzaketa
Aurreko sagu gurutzaketan, alelo gainartzaileak kromosoma batean baldin badaude, eta atzerakoiak alelo homologoan, badagogaldetzerik zergatik ateratzen diren F
2
an
alelo gainartzaile eta atzerakoi baten arteko
konbinazio berriak. Badirudi F
2
batzuetan
elkarrekin lotutako lociek lortu dutela
banatzea, eta independentzia osoz konbinatu
direla gero berriz elkarrekin. Oso ondo
ikusten da hori F
1
sagu batzuekin frogazko
atzeragurutzaketa egiten denean. Ikus dezagun
agertu ohi diren emaitzak:Horren ondorioz, praktikan ez dago erabateko
loturarik, eta ager daitezke forma
berri batzuk, gurasoenak ez bezalakoak, berkonbinagai
deritzaienak. Horren zergatia,
berriz ere, meiosian dago. I profasean, kromosoma
homologoek pareak sortzen dituzte,
kiasmak osatzen eta puskak, zein bere
kromosomarekin, truka ditzakete eta, hala,
trukegurutzaketa sortu. Era horretan, II
anafasean alelo konbinazio berriak sortzen
dira. Horren ondorioz, kromosoma baten
loci bateko A aleloa, zelula zenbaiten meiosian
zehar, B alelotik bana daiteke, kromosoma
beraren beste loci batean baitago hori,
eta kromosoma homologoan dagoen azken
horren b aleloarekin elkartu.
Independentziaren legea, beraz, aski fenomeno
nahasia da. Zenbait kasutan erabatekoa
izaten da independentzia. Besteetan, dependentzia
edo lotura izaten da. Lotura, bestalde,
partziala izan ohi da, maila askotakoa.
Zeren arabera gertatzen da bi ezaugarriren
arteko lotura maila handiagoa edo txikiagoa
izatea? Batez ere, meiosian zehar beren geneen
locien artean gertatzen diren trukeguratzaketa
kopuruaren arabera. Trukeguratzaketak
ez dira nahi eta ez gertatzen meiosian
dauden zelulen %100ean; zoriaren arabera
gertatzen dena. Zenbat eta urrunago egon bi
lociak, handiagoa izango da trukegurutzaketak
gertatzeko probabilitea. Distantzia da
hor erabakigarria. Zenbat eta handiagoa
izan distantzia, handiagoa da trukeguratzaketen
portzentaia, eta, horren ondorioz,
handiagoa izaten da forma berkonbinagaien
maiztasuna. Ondorio gisa esan daiteke bi
generen arteko berkonbinazio maiztasuna
beren locien arteko distantzia fisikoaren
neurri gisa erabil daitekeela.
Maiztasun hori ondorengo formula hau
erabiliz kalkulatzen da, atzerako gurutzaketa
bati aplikaturik:Lehen aipatu den esperimentuaren kasuan
ikusten denez, %10 berkonbinagai atera dira.
Balioak izan daitezke, ia %50etik hasita, hori
kromosomaren ertzetan kokatuak baldin
badaude bi geneak, %0raino, iai-ia elkarri
itsasita badaude. Proportzioa %50 denean,
Mendelen legeko independentziaren edo
lotura falta erabatekoaren baliokidea da, hau
da, kromosoma banatan daude bi geneak.

Geneen mapak
Ikusi berri dugu ezen, bi gene kromosoma
berean daudenean, meiosiaren unean gene
233horien artean gertatzen diren trukegurutzaketen
proportzioa elkarren arteko distantziaren
araberakoa dela. Trukegurutzaketa
geneen arteko distantziaren neurria baldin
bada, baliagarri izan daiteke, logikoa denez,
geneek kromosoman duten kokalekua, besterik
ez bada erlatiboa, ezagutzeko. Hori
dela-eta, geneen kokaleku erlatiboak erakusten
dituzten mapak egin ahal izan dira.
Mapa banakoa (km genetikoa bezalako zerbait
balitz bezala), berkonbinazioen %1 gertatzen
den bi locien arteko distantzia gisa
definitu izan da. Izena ere eman zaio banako
horri, zertimorgana.
Morganen genetista taldea, Estatu Batuetan,
nabarmendu da gehien genetikaren
alorrean. Zientzialari hauen hipotesiaren
arabera, geneak bata bestearen atzean daudekokatuak kromosoman, hau da, lerroan
ipiniak; frutaren euliaren (Drosophila melanogaster)
kromosomen mapa genetikoak egitea
lortu zuten. Hain zuzen ere, zientzialarihonen ohorez asmatu zen zentimorgan
izena.
Adibide erraz batek lagunduko digu ulertzen
nola egin daitezkeen mapa hauek:
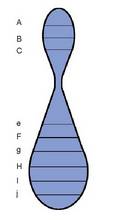
Eman dezagun badirela hiru gene, A, B eta
C, elkarrekin lotuak, hau da, kromosoma
berari dagozkiola. Gurutzaketak egiten dira,
haien artean gertatzen diren berkonbinazioak
kalkulatu ahal izateko. Ondorengo hauek
dira: A-B=%30; A-C=%12; B-C=%42
Kromosoma irudikatzen duen lerro bat
marraztuta, hau egin genezake:
1) Erdian jarriko ditugu elkarrengandik
hurbilen dauden geneak, A-C=12; hurrengo
distantzia hartu, A-B=30, eta jarriko dugu B
bi kokaleku posibleetan; 3) gauza bera egingo
dugu azkenekoan, B-C=42. Horrekin B
ezkerrean ipini genezake.Baina hain erraza ere ez da izaten mapak
egitea. Asko zailtzen duten eragozpenak
sortzen dira, hala bi geneen arteko trukegurutzaketa
bikoitzak; fenomeno hori oso gertagarria
da geneak aski urrun daudenean;
kromosoma bateko parte desberdinek duten
trukegurutzaketa maiztasun desberdina izaten
da beste arazo bat. Hortaz, berkonbinazio
maiztasunen bidez locien kokaleku erlatiboak
jakin daitezke, baina ez dira
fidagarriak izaten beren arteko distantzia
erreala finkatzeko. Horrez gainera, jakin
beharko litzateke zehatz zein kromosomatan
dagoen gene talde bakoitza, eta orobat gene
bakoitzaren kokaleku zehatza. Horretarako,
bestelako fenomenoak aztertu behar izan
dira (kromosoma aberrazioak, kromosoma
zerrendatzea, etab.), baina horiek ez dira
hemen landuko.
Zailtasun horiek hobeto ulertzeko, pentsa
dezagun mapa bat eman digutela; mapa
horretan oso ondo agertzen dira hainbat hiri
errepide batean ertzean kokatuak, eta orobat
horien arteko distantziak ere, kilometrotan.
Baina mapa zati horretan bestelako informaziorik
ez dagoenez, eta hiri horiek guztiz
ezezagunak ditugunez, galdegin daiteke ea
izango ote dugun jakiterik zein herrialdetan
dauden hiriak, eta, herri horretan, zein partetan
dauden.
Loci desberdinen arteko berkonbinazioaren
maiztasunaren hastapen honi esker, mota
askoren mapa genetikoak ezagutu ahal izan
dira, baita gurea ere. Giza genetikaren atalean
esaten denez, gure motaren kasuan zailtasun
handiak dira kromosomen mapa genetikoak
egin ahal izateko. Nolanahi ere,
aurrerapen handiak izan dira, eta, hala, X
kromosoman bakarrik, 350 loci baino gehiago
ezagutzen dira. Genetika molekularraren
teknika modernoen aplikazioak garrantzi
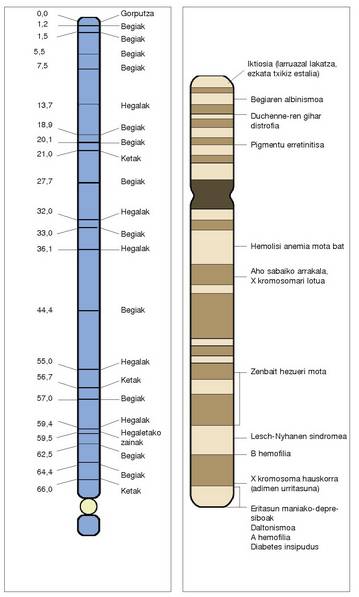
handiko emaitzak eman ditu alor honetan.Mapa horien laguntzaz jakin genezake,
adibidez, ABO odol taldeen genea 9 kromosoman
dagoela, eta, besteak beste, ondoko
ezaugarrion erantzule diren geneekin lotua
dagoela: azkazal-errotula sindromea; dentsitate
apaleko lipoproteinen hartzailea (kolesterolaren
kontrola); hainbat interferoi. Eraberean, Rharen genea 1 kromosoman ipini
genezake, amilasen eta Duffy odol taldeen
(hori da autosometan kokatu ahal izan zen
lehen locusa) geneei loturik. Hori jakitea
oso garrantzitsua izan da sendagintzan,
herentziazko eritasun askoren ezagutza
bidea eman baitu.